
7. The Mutual Transcription of Metaphors in the Fields of Natural Sciences, Social Sciences, and Cultural Studies
1 2018-02-27T13:26:13-08:00 Norman Salusa cd154f315b608a99b35584de6a697dca17b7e6af 21206 1 plain 2018-02-27T13:26:13-08:00 Norman Salusa cd154f315b608a99b35584de6a697dca17b7e6afThis page is referenced by:
-
1
2018-02-27T13:21:53-08:00
7. The Mutual Transcription of Metaphors in the Fields of Natural Sciences, Social Sciences, and Cultural Studies
38
7. The Mutual Transcription of Metaphors in the Fields of Natural Sciences, Social Sciences, and Cultural Studies
plain
2018-06-14T08:53:43-07:00
The draft of a theory and empirical analysis of religious evolution programmatially presented here is, among others, based on approaches in natural sciences. As was suggested, this is possible due to a isomorphism between religious, social, mental, organic and physical evolution. However, the possibility alone does not tell us anything about its usefulness. The purpose of this transfer is to bring the study of religion to areas beyond other-referential elements of hermeneutics (i. e. interpretation that is oriented towards persons and the subjectivities), to scientify it, and to center it in a self-referential way. This approach is meant in the sense of a “third culture” (Brockman 1995; Lee and Wallerstein 2004), which overcomes the dichotomy of the “two cultures” (Snow [1959] 2012) and which brings cultural studies, the social sciences and the natural sciences together to form a unified science. (1) Religion is no more a hermeneutic fact than ‘nature’—at least as an object of communication—is exclusively bound to ‘hard facts’.
Against this background, the scientific description (i. e. ‘deciphering’) of the genetic code and cell processes is transferred to the description of semiotic processes in order to inform the study of religion. (2) It is also in the use of the concept of the meme that this must be done consistently if metaphorics is to hold up heuristically (Lynch 1996; Blackmore 1999; Aunger 2000; Blackmore 2001; Distin 2005; Dawkins 2006; Brodie 2009; Heylighen and Chielens 2009; Leigh 2010; Blackmore 2016). (3) The general semiotic code, which is responsible for the composition of a complete sign, is interpreted in this approach in analogy to the biogenetic code and corresponding cell processes—just as, vice versa, the cell can be interpreted as a semiotic system (Barbieri 2007b). (4)
Signs are as aggressive as genes and they push for reproductive and mutating development. (5) On the same basis as the general semiotic code, they each form specific semiotic codes—for example, a religious one—, transmit themselves by means of sign division and combine to form sign systems that are organized on the basis of specific semiotic codes. (6) “Cultural transmission is analogous to genetic transmission” (Dawkins 2006, 189). But the analogization of biogenetic and semiotic code—at least in the approach advocated here—does not involve a naturalized understanding of socio-cultural reality and therefore also of religion, namely for two reasons. First, an analogy does denote similarity in some respect (i. e. similarity in difference to differences), but not identity. Second, the biogenetic code itself is a metaphor of prebiotic chemistry and molecular biology (7) borrowed from the subject of scripture and its deciphering (Brandt 2004; Stoschus 2005; Türcke 2005). (8) Insofar as the structural sciences are oriented by the reference medium of language and writing, they “increasingly contribute to the ‘dematerialization’ of the natural sciences, which in turn makes it possible to bridge the gap to the humanities” (Küppers 1996, 197). Scripture and the ability to read it are absolute metaphors as conceptualized by Hans Blumenberg (2010). They characterize not only cultural studies and social sciences, but also the natural sciences. (9) In accordance with the mutual analogization of the biogenetic and the semiotic codes, the semiotic structure of communication, as well as the biogenetic code, are degenerated, universally (10), and as a whole, they cannot be put at rest. Just like communication, the biogenetic code has in information-theoretical, syntactic and semantic terms a demonstrably high error tolerance and adaptability (Natterer 2010, 92; cf. also Mattick 2003). (11) By analogy with the relationship between microphysical and macrophysical processes (12) as well as with processes in a cell and the numerous forms of organisms, the complete sign constitutes the elementary process of communication, which takes the form of semantic textures (13); in terms of societal structure it becomes a societal system with differentiated subsystems, and in the social morphogenesis it takes the form of different social forms—ranging from the interaction in the form of an encounter (Kieserling 1999) to formal organizations (Luhmann 2000). This is the empirical-sociological correlation between semantics and social structure. The semiotic code contains the arrangement of the mechanisms that are essential for communication and it unfolds in discourses as well as in fixed socio-structural arrangements. Socio-cultural evolution, the autonomous part of which is the evolution of religion, is therefore not to be interpreted as an extension of biological evolution—that would be an incorrect form of reductionism—but once again: in the sense of analogy to it. (14) As was noted earlier, biological and socio-cultural evolution form a relationship of a structural isomorphism. In the following passages, this will be explained on the basis of analogies between semiotic processes and cell processes.
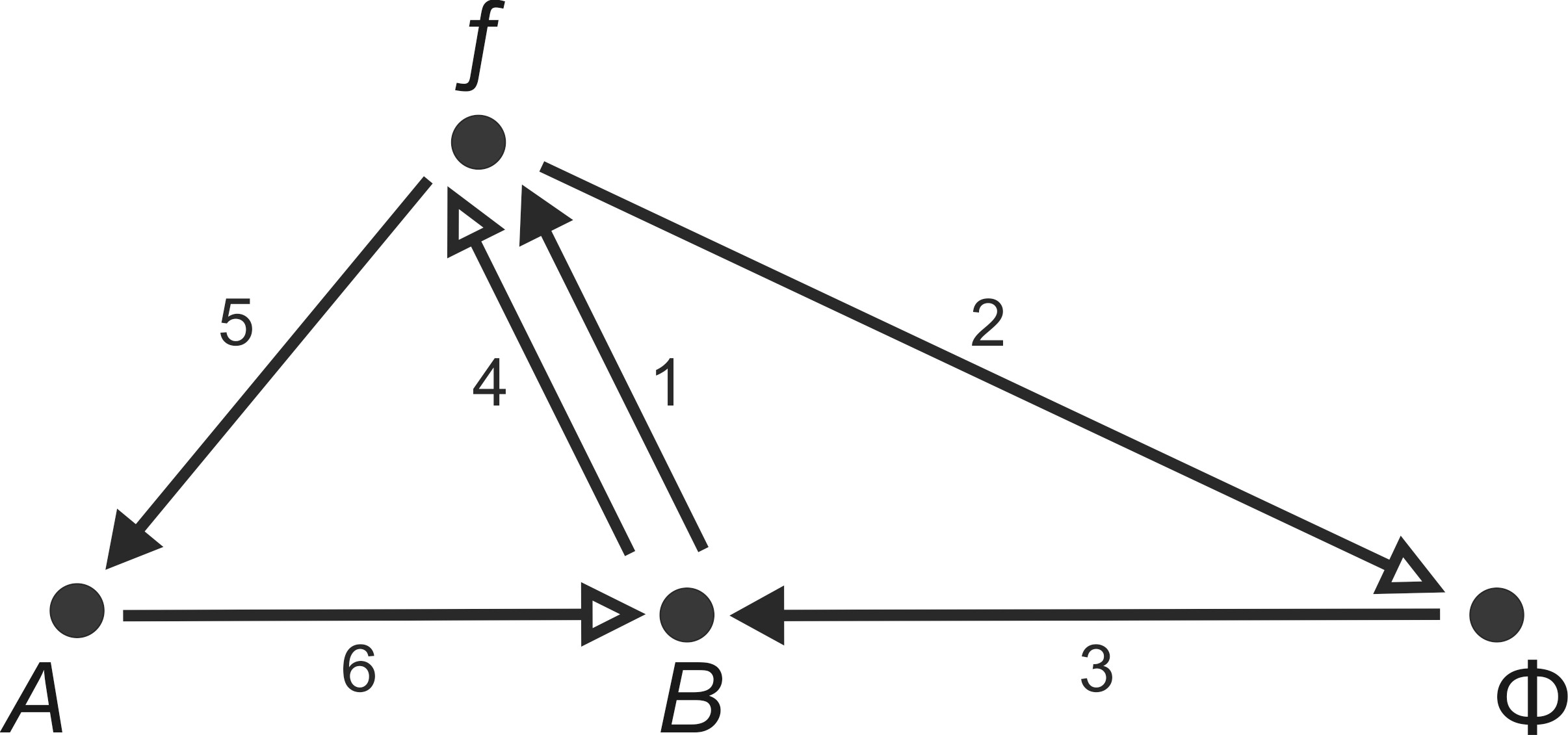
I am far from understanding cell biology and its theory models. However, a formal analogy between the corresponding diagram and the semiotic model developed here quickly springs to one’s mind. The theoretical biologist Robert Rosen modeled the so-called (M,R)-system, where ‘M’ stands for metabolism and ‘R’ for repair (15) (Rosen 1991). (16) The diagram looks like this (Figure 14):
(in accordance with Rosen 1991, 251) (17)
The model is composed of the following processors and processes (also known as functors and functions, or operators and operations): the metabolical processor ƒ uses inputs from set A to produce outputs of set B. ƒ is caused by the repair processor Φ, which in turn is generated by a replication processor b from set B. The lines with hollow arrowheads stand for “an actual software flow from input (afferent) to output (efferent)”, whereas the lines with solid arrowheads stand for “the induction or generation of this flow by the hardware” (Rosen 1991, 219–220). (18) The result is a circular system: “[...] every function is indeed entailed by another function in the graph itself. As far as entailment is concerned, the environment is out of the picture completely, except for the initial input A” (Rosen 1991, 251). The relations are (see also Cottam, Ranson, and Vounckx 2007, 2358):
ƒ→a→b (metabolism; relations 5 and 6)
ƒ→ϕ→b (repair [or exchange] function; relations 2 and 3)
b→ƒ (replication [or closure] function; relations 1 and 40 (19)
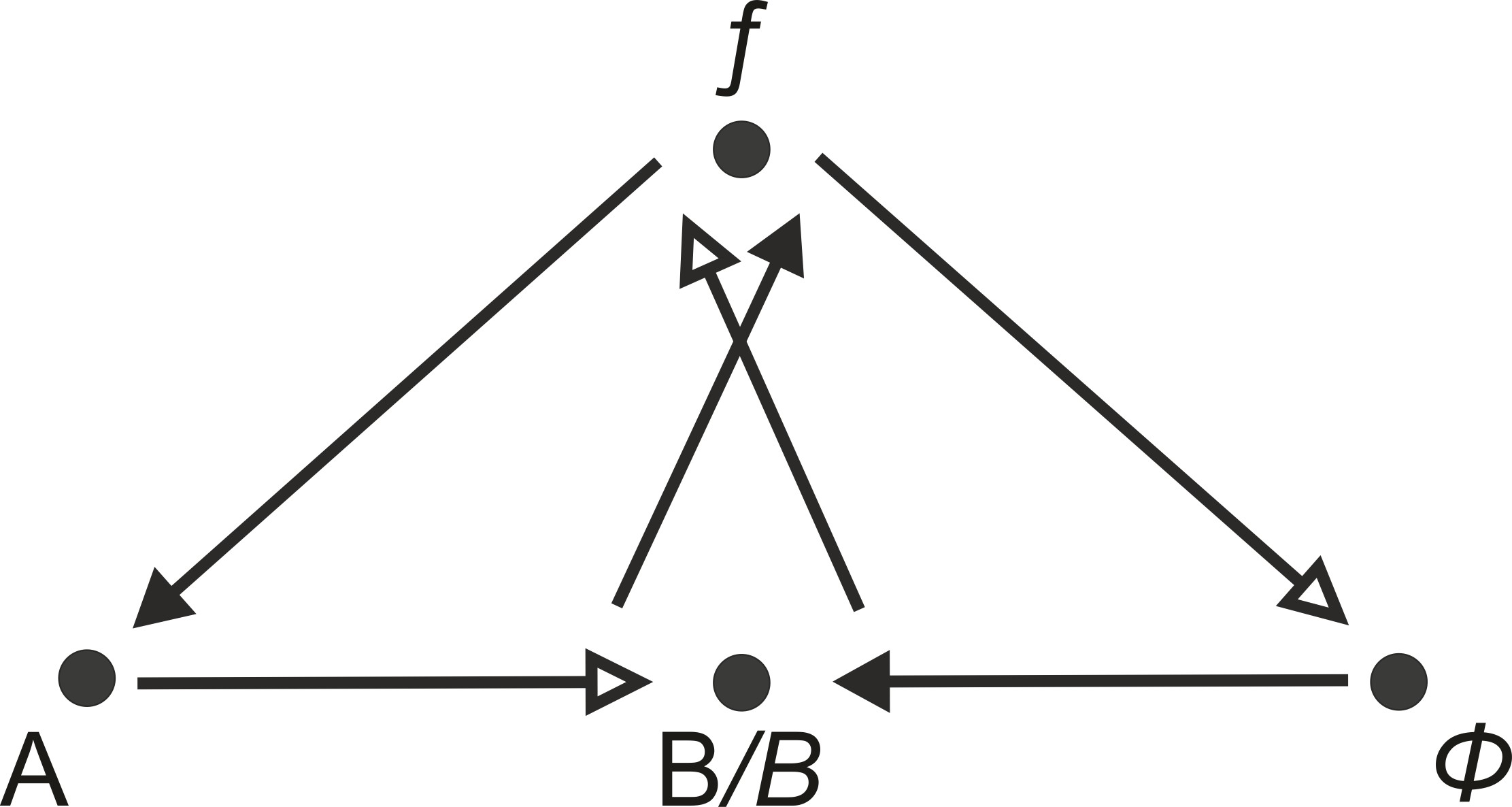
However, this model is associated with a number of difficulties—especially with regard to the relationship between mechanism (hardware) and organism (software). (20) Although in the (M,R)-system a mechanism can ‘contain’ an organism and an organism a mechanism, this distinction remains sharply binary. “Abstractly, the recognition in general of a single object implies the existence of not two, but three separate domains: the object, its ecosystemic environment, and their interface: the bifurcating categorization of Nature proposed by Rosen into the complement of mechanism and organism is insufficient. An organism is not the complement of a mechanism: the complement of a mechanism is its ecosystem” (Cottam, Ranson, and Vounckx 2007, 2359). The latest research suggests a complex interplay between digital and analog processes within the cell: “[…] it now seems likely that a major part of reproductive and embryogenetic control is not exercised by the small percentage of DNA that, seemingly directly and digitally, controls protein synthesis. Many of the other DNA-related processes now coming to light depend on a multiplicity of more or less locally environmental analogue effects. This ‘insertion’ of analogue influences into the previously supposedly digital transcription-synthesis route between gene and protein has enormous consequences. A prime attribute of digital or quantized interactions is their insensitivity to ‘noise’ or small-scale, locally environmental influences. This simplifying isolation breaks down when analogue effects come into play, leaving the door wide open to genetic influence from the environment. [...] Chemico-physical epigenetic codes are far more susceptible to environmental influence than are genetic codes” (Cottam, Ranson, and Vounckx 2007, 2357). Accordingly, Cottam, Ranson, and Vounckx (2007, 2360) see relations 2 and 3 in the (M,R)-system (Figure 14, above) as the ecosystem which complements the mechanism. An organism is, therefore, “the complex interface between mechanism and ecosystem” (Cottam, Ranson, and Vounckx 2007, 2363). The ecosystem corresponds to the transition between the non-specific and the specific environment of a system, as was described in the chapter on the foundations of systems theory (see Figure 1). The diagrammatic solution offered by the authors is as follows (Figure 15): (21)
(in accordance with Cottam, Ranson, and Vounckx 2007, 2358)
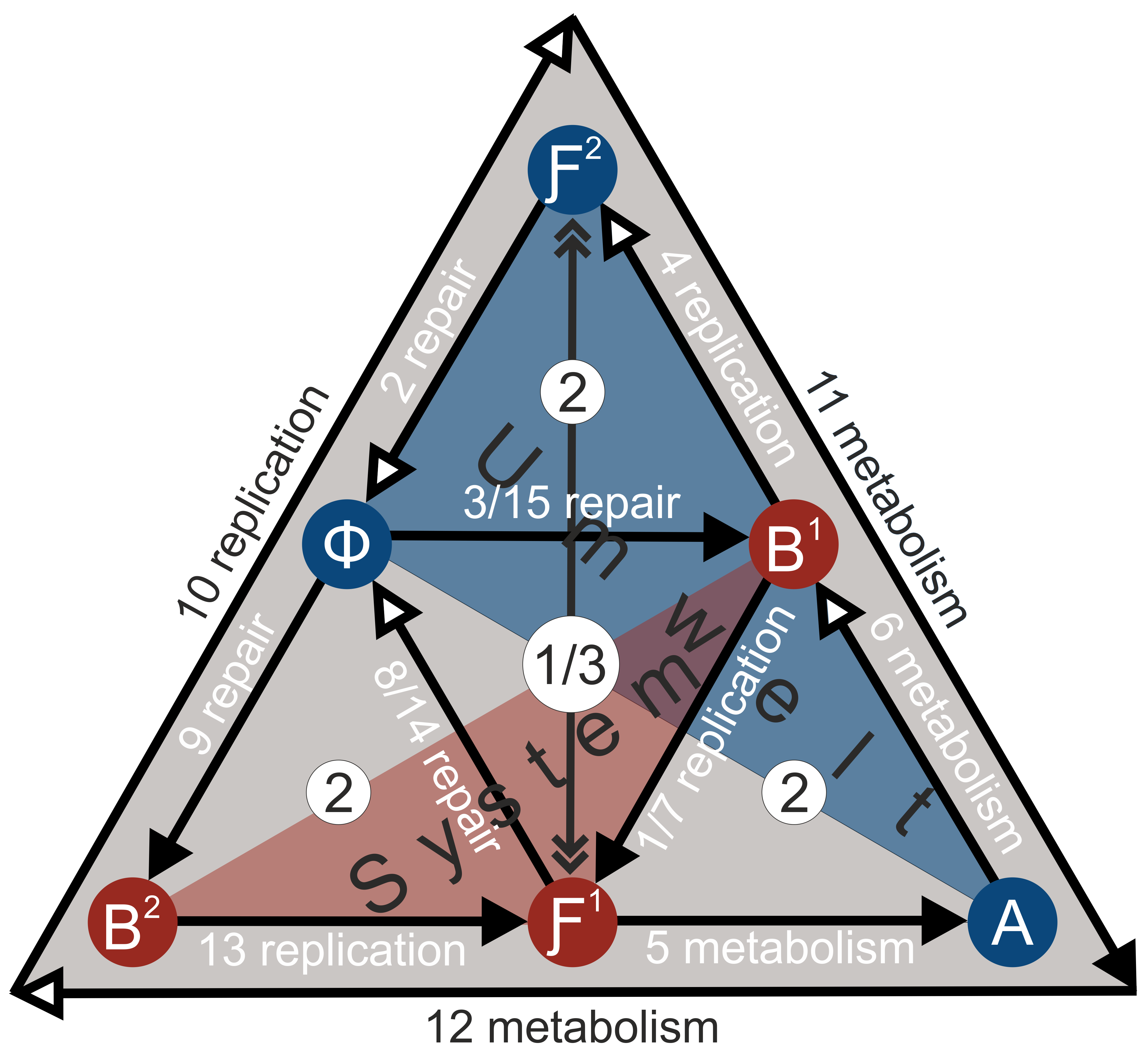
If the (M,R)-system is autopoietic and self-referential, the functions not only have to be caused by one another, but there must also exist foundations of the relations (Goudsmit 2007). If one refers the (M,R)-system to the considerations on the double input-output process, which were discussed in the chapter on the foundations of systems theory (cf. Figure 1), as well as to the resulting considerations on the semiotic elementary system (Figure 7) then the diagram of the (M,R)-system is to be modified as follows:From a semiotic self-referential perspective, the following applies: “[…] self-reference on the level of basal processes is possible only if at least two processing units which operate with information are present, and only if they can relate to each other and thereby to themselves” (Luhmann 1995c, 138). In addition, the system must go through the function B→ Ƒ four times, through the function Ƒ→Φ→B three times, and through the function Ƒ→A→B two times in order to reach a closed state. The fully developed system starts with function 1 (B1→Ƒ1) and ends with function 15 (Φ→B1) (Figure 16). Circularity creates a “bizarre hierarchy” of three levels, each with its own elements, “in which it is the case that, for each of the three members, that particular member is exactly in the middle of the hierarchy, is bizarre (counter-intuitive) but non-absurd (logically non-contradictory)” (Kercel 2002, 135). (22) The replication function B2→Ƒ2 (function 10) and the metabolism Ƒ2→A→B2 (functions 11 and 12) constitute a third level and fold in the preceding processes of the second and first level.
This modeling of the second-order would have to be—if it turns out to be useful—further considered, elaborated and, if necessary, modified further by experts, which cannot be done here. (23) Perhaps something for biosemiotics has been gained with it; I cannot verify that. But even if this modeling cannot be maintained from the point of view of theoretical biology, the analogization is still heuristically helpful for the understanding of the semiosis of socio-cultural processes in general and thus also for religion in particular. Then it could be called a productive misunderstanding. By analogy with the arrangement of the (M,R)-system in accordance with the semiotic model in the operatively closed state (Figure 16), the following arrangement of the sign elements in semiosis can be made:
Analogization makes the following points clear:
- The sign system creates an ‘image’ of itself and its environment within the system. This is achieved by observing the first inner level through the third outer level.
- The middle level of semiosis is characterized by the system-internal interplay between systemic code and environmental influences. Therefore, it represents the distinction between the observer and the observed, unified by the third level. The middle level corresponds to Stanley Salthes’ conception of the “focal level”, which is positioned between the lower and higher levels—or, according to the folding metaphor of the semiotic model: between the inner and outer levels (see Salthe 1985, esp. 125). (24) The middle level can only be observed by an external observer (Salthe 1985, 290). Due to the interplay between systemic code and environmental influences, the direction of the process reverses at the middle level. In the flowing state of the system, this is possible between the higher (or outer) and lower (or inner) levels: “[…] an entity can be both a and not-a because it is in the process of changing away from being a. A system of this kind would be indeterminate because it is in flux” Salthe 1993, 232, referring to Voorhees 1985). However, systems must be studied when they are in a synchronous state, “because it is only then that they gain a quasi-determinacy sufficient to characterize them” (Salthe 1993, 232, referring to Voorhees 1983).
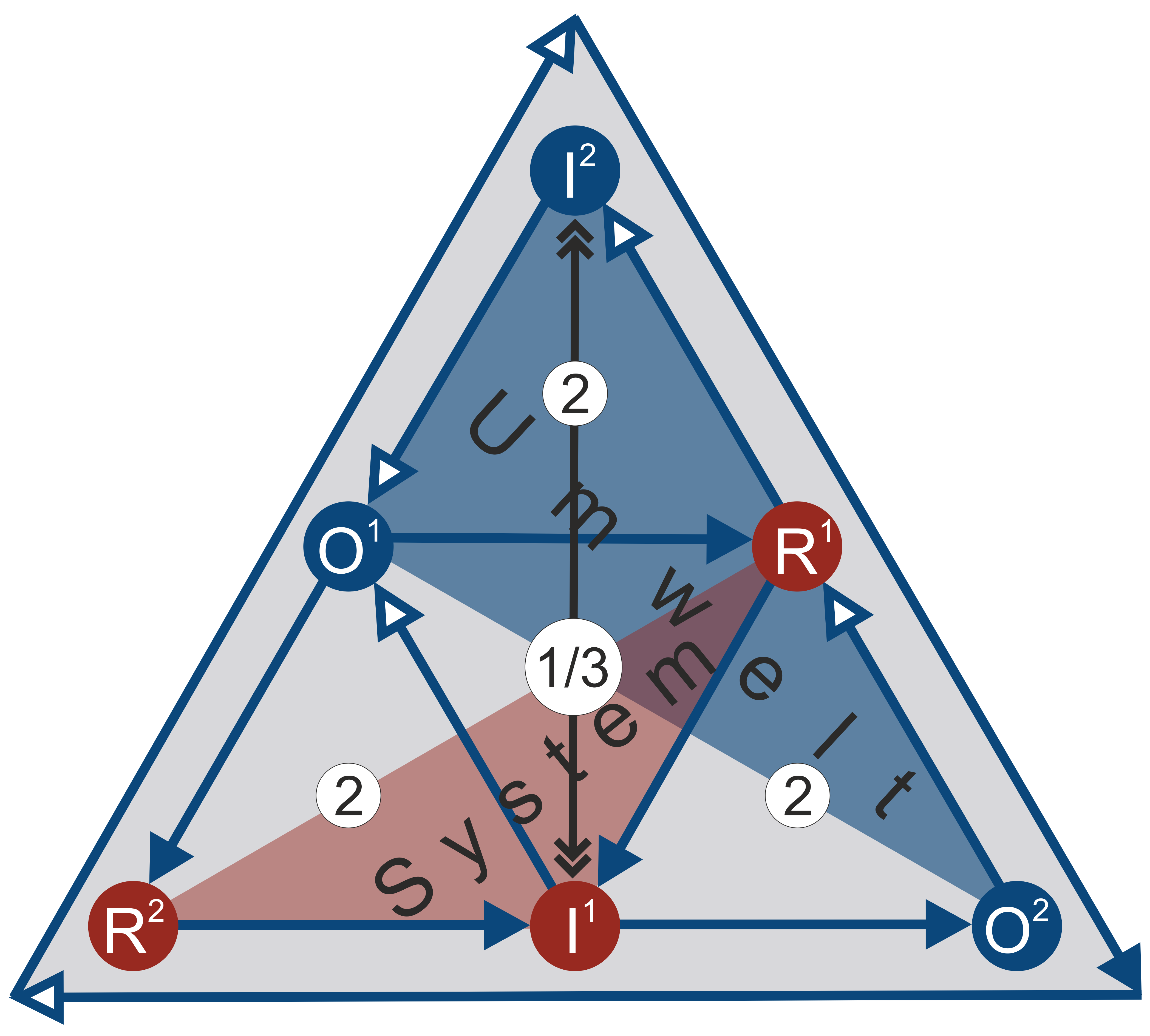
Against this backdrop, the relationship between metonymy and metaphor discussed above (cf. section 6.2) should be reexamined. (25) The analogization between cell processes and socio-cultural semiosis results in the following correlations:
- The two metaphorical representamen R1 and R2 correspond to output processors B1 and B2.
- The metonymic sign object O1 corresponds to repair or replacement processor Φ.
- The metonymic sign object O2 corresponds to input processor A.
- Interpretants I1 and I2 correspond to processors Ƒ1 and Ƒ2 of the (M,R)-system. They are responsible for replicating the semiotic code. Due to the pragmatic environmental sensitivity, I2 additionally ensures that environmental influences are taken into account.
- The syntagmatic functions R1→I1 and R2→I2 are key to the semiotic code. They correspond to the replicating (or closing) functions B1→Ƒ1 and B2→Ƒ2 and render a self-referential foundation of the sign system possible.
- The semiotic function I2→O1→B1 corresponds to the repair (or exchange) function Ƒ2→Φ→B1 and thus to the ecosystem described by Cottam, Ranson, and Vounckx (2007) as the complement to the mechanism of the (M,R)-system.
- The semiotic function I2→O2→R2 corresponds to the metabolic function Ƒ2→A→B2.
In keeping with the analogy between cell processes and communicatively activated semiosis, it is possible to understand how the religious system draws semantic energy from its environment and transforms it into system-specific information (Figure 17): It starts at the representamen R1 as a unity of system and environment. Its metaphorical and ambiguous content is transformed by the interpretant I1 as one of the system processors into clear, unequivocal religious information. In this way, the religious code is replicated. The interpretant I2 as the second system processor then draws semantic energy from the specific system environment—or, mediated through it, from the non-specific environment—in the form of the sign object O1, which is coordinated with the first representamen R1. This is an analogy to the repair (or exchange) function of the (M,R)-system and at the same time the environmental sensitivity of the semiotic system in terms of its “ecosystem” (Cottam, Ranson, and Vounckx 2007, 2358–2359). Furthermore, the system draws semantic energy again from the system-specific environment in the form of O2 and transfers it—mediated via interpretant I2—into system-specific information in the form of the representamen R2. R2 is partially released into the system environment and partially constitutes the starting point for the replication of the religious code (B1→I1). In other words, R2 is treated as a metaphor, the polysemy of which is then divided up into environment and system-specific information by means of a bifurcation.
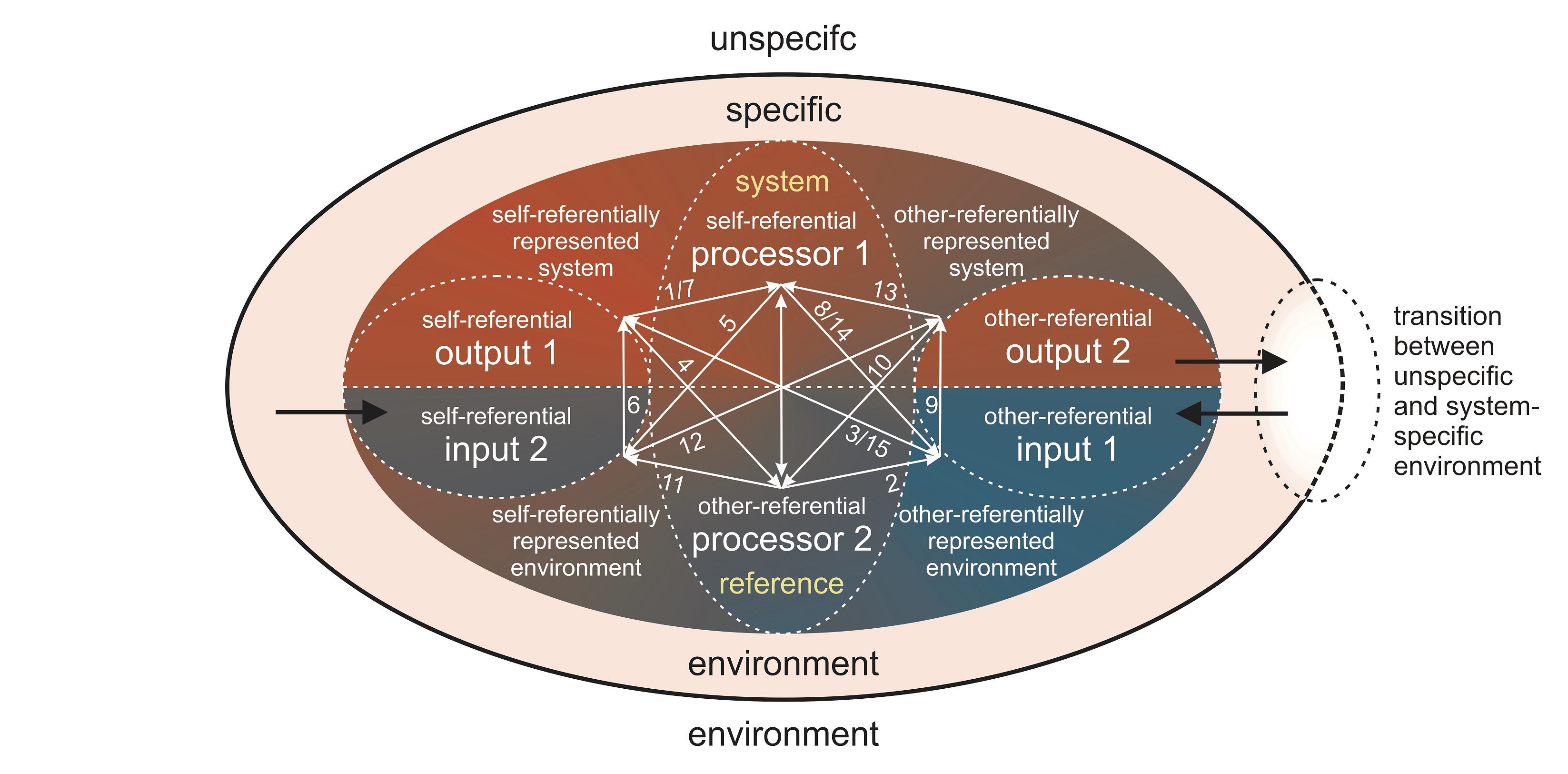
Based on the mutual analogization of the (M,R)-system and the elementary sign system, it is also possible to specify the double input-output process (Figure 18), which was already the subject of the chapter on systems theory.
This way it becomes apparent how self-reference, other-reference, and system-reference interact with each other. The autopoietic process starts with the self-referential output 1, where the self-referential processor 1 has its starting point (relation 1). From there, the process takes the path to the other-referential processor 2, and, via the other-referential input 1 and the self-referential output 1, back to processor 2 (relations 2–4). The process then switches to the self-referential processor 1, which triggers the self-referential input 2 and output 1 (relations 5 and 6). Output 1 once again stimulates processor 1 (relation 7), which then regulates the other-referential input 1 and output 2 (relations 8 and 9). Output 2 stimulates processor 2 (relation 10), which controls the self-referential input 2 and the other-referential output 2 (relations 11 and 12). Output 2 stimulates processor 1 (relation 13), which eventually regulates other-referential input 1 and self-referential output 1 (relation 14 and 15).
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}